石油分解菌の検出とその多様性
石油分解菌の群集構造
本調査では、まず、実際の自然海水中にはどのような細菌が生育しているかを、変性剤濃度勾配ゲル電気泳動(DGGE: Denaturing gradient gel electrophoresis)を用いて解析しました。次いで、この自然海水に原油及び栄養塩を加えることによって、微生物の群集構造がどのように変化するかを解析しました。最後に、環境中において石油分解に関与する細菌群を明らかとするために、原油及び栄養塩を添加した海水中で石油分解菌群を増殖させ、そこに含まれる石油分解菌の種類を比較解析しました。
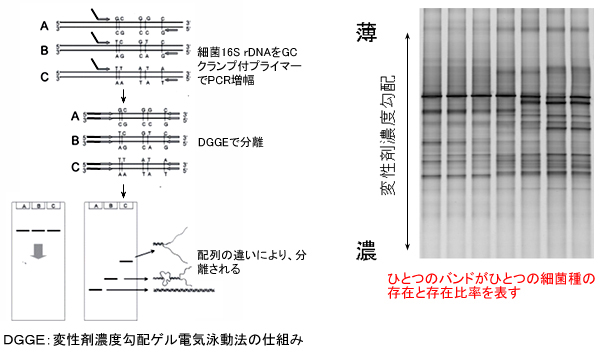
採取前に現場の海水で十分に洗浄した20リットル容のポリプロピレン製タンクに海水を採取し、海水中の微生物は孔径0.22μmのメンブレン・フィルター上に捕集しました。捕集済みのメンブレンはドライアイスで凍結、または4℃で保冷して研究室まで輸送しました。このフィルターから、ビーズビーティング法を用いてDNAを抽出しました。次いで、このDNAから細菌16S rRNA遺伝子断片をPCR増幅しました。細菌16S rRNA遺伝子断片のPCR増幅にはGCクランプを付加したプライマーを用いました。このようにして得られた16S rRNA遺伝子断片について、DGGEを用いて微生物群集構造を解析しました。DGGEでは、DNAの変性の度合いの違い(一本鎖DNAへの解離しやすさ)を用いて、DNA断片を分離します。DGGEゲル上のバンドそれぞれは、異なった細菌から由来した16S rRNA遺伝子断片と考えられます。主要なバンドの塩基配列を決定し、得られた塩基配列はBLASTにより相同性検索を行いました(【図1】)。
【図1】細菌群集構造の解析方法 / DGGE:変性剤濃度勾配ゲル電気泳動法
自然海水中の細菌群
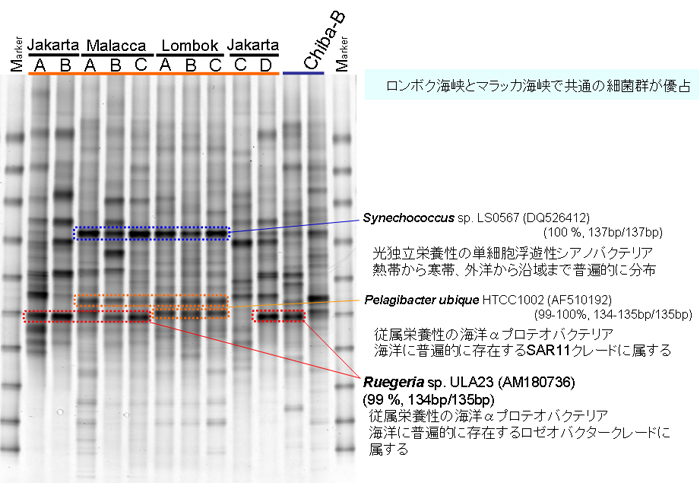
原油で汚染される前の自然海水中の細菌群のDGGEプロファイルを【図2】に示します。マラッカ海峡及びロンボク海峡の全地点において、共通してSynechococcus属及びPelagibacter属が優占種として検出されました。また、日本沿岸とインドネシア沿岸において共通した優占種としてRuegeria属が検出されました。マラッカ海峡とロンボク海峡においては、外洋からの一定の海流が存在することから、陸地からの排水の影響等が少なく、両者の細菌群集構造が類似していると考えられます。
【図2】DGGE による自然海水中の菌相解析
原油・栄養塩添加後の石油分解菌群
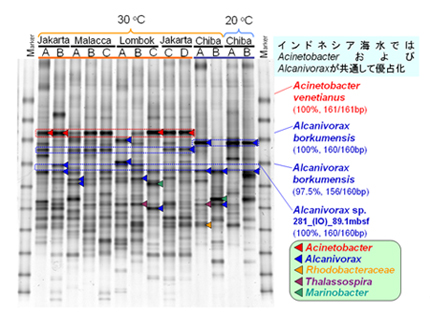
清浄な海水中には、微生物を高い密度にまで増殖させるのに必要な栄養塩(主に窒素とリン)が十分量含まれていません。そのため、原油で汚染された海水にこれら不足している栄養塩を加えることによって、石油炭化水素を炭素源として利用できる微生物の増殖を促進させ、原油の分解を加速することができます。このように、微生物を用いて環境汚染を浄化する技術を一般にバイオレメディエーションと呼びます。その中で、現場に生育する石油分解菌の増殖を促すことによって汚染を浄化する技術をバイオスティミュレーションと呼びます。一方、汚染現場に微生物を散布することによって汚染を浄化する技術をバイオオーグメンテーションと呼びます。原油・栄養塩添加後の石油分解菌群の比較を行いました。原油と栄養塩を加えて10日間30℃で振とう培養した後の培養液中の細菌群集構造をDGGEにより解析しました。ロンボク海峡A地点(【図3】)を除く全地点でAcinetobacter venetianusが最も優占化していました。日本沿岸海水においては、Alcanivorax borkumensisが最も優占化していました。インドネシア沿岸海水においても数地点においてAlcanivorax borkumensisが検出されましたが、日本沿岸海水のバンドとは移動度が異なり、日本のAlcanivoraxとは異なる株であると考えられます(【図3】)。
【図3】原油と栄養塩添加後の細菌群集
石油分解菌群の時期的変動
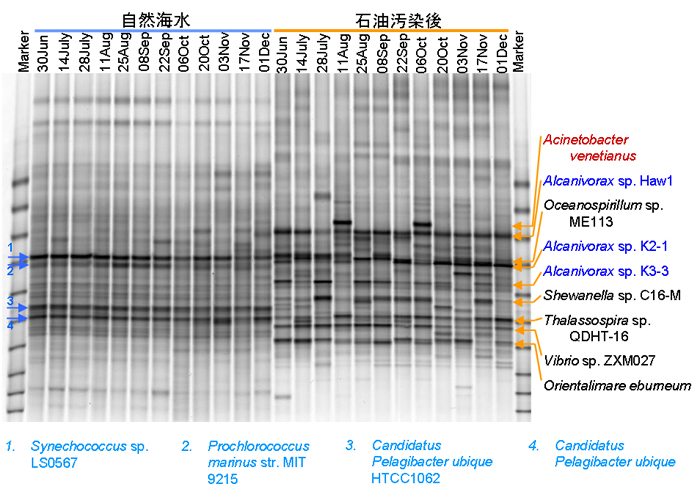
インドネシア沿岸海水中の細菌群及び石油汚染時にインドネシア海水中に優占化する細菌群について、時期的な変動を解析しました。海水はロンボク海峡B地点(【図4】)において、2008年6月30日から同年12月1日まで2週間ごとにサンプリングしました。この自然海水に栄養塩と原油を加え、10日間30℃で振とう培養しました。これら自然海水及び原油分解バッチ培養中の細菌群集構造をDGGEにより解析しました。その結果、自然海水中では、実験期間を通して細菌群集はほぼ一定であり、Synechococcus属及びPelagibacter属が優占種として検出されました。原油添加区においても、実験期間を通してほぼ一定の細菌群集を示し、2種のAlcanivorax sp.が優占種として検出されました。インドネシア各地の沿岸海水中で石油汚染後に優占化したAcinetobacter venetianusは2サンプリング時点(8月11日と10月6日)でのみ、優占種として検出されました(【図4】)。【図3】に示した地点間の比較結果においても、ロンボク海峡B地点におけるAcinetobacter venetianusの優占化度は低く、同地点においては、主としてAlcanivorax属が初期の石油炭化水素分解に寄与することが推察されました。
【図4】石油分解菌の分離
以上をまとめると、以下のようになります。
- 栄養塩の添加により石油分解菌が増殖し、石油分解が促進されました。
- インドネシアと日本では、自然海水中及び石油汚染後に増殖する石油分解菌群の構成が異なりました。
- インドネシア海域における石油汚染に対して栄養塩添加(バイオスティミュレーション)が有効であり、熱帯海域における活性化手法の開発が必要です。
インドネシア海水からの石油分解菌の分離
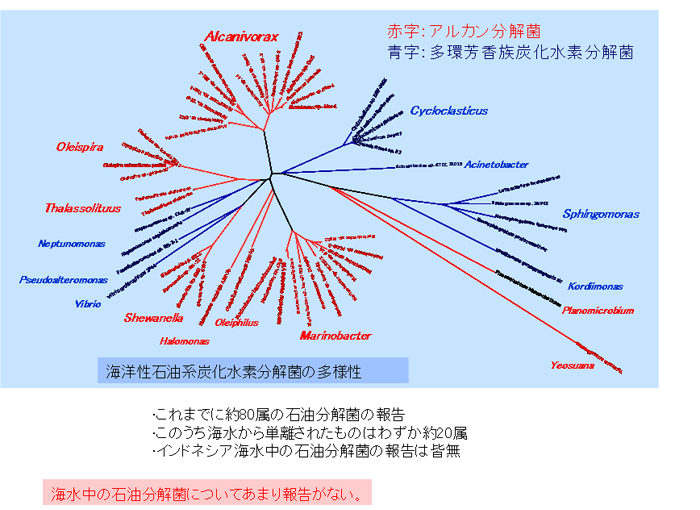
海洋性の石油系炭化水素分解細菌に関する研究は60年以上前から行われていますが、海洋からは、これまでに19属の細菌しか分離されておらず、陸域から分離されている約80属に比べわずか4分の1程度です。さらに、分解細菌はこれまでに主に冷帯・温帯域から分離されていることが多く、インドネシアのような平均海水温が30℃の熱帯海域にどのような分解細菌が存在するのかについてはほとんど知られていません(【図5】)。
【図5】既報の海洋性石油分解菌
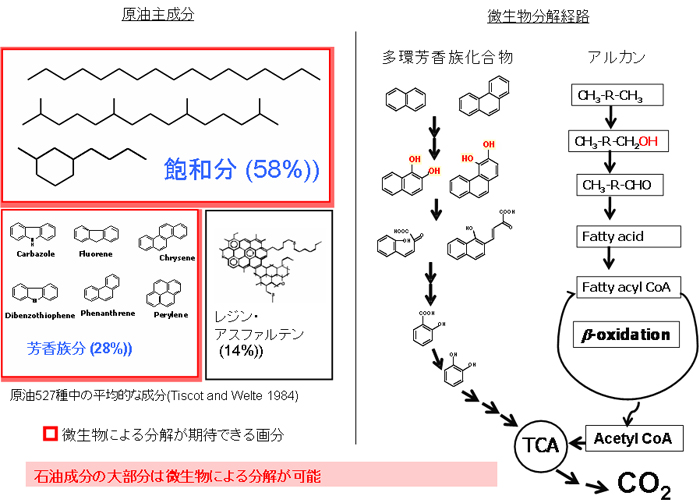
- 参考:原油成分の微生物分解【図6】
- 原油には、非常に多くの種類の炭化水素及び、S、N、Oを含む炭化水素誘導体が含まれます。これらは、一般に脂肪族分(飽和分)、芳香族分、レジン分、アスファルテン分に大分類されます。飽和分は石油の主成分であり、n-アルカン、分岐鎖アルカン、環状アルカン等から構成されます。芳香族炭化水素としては、ベンゼン、トルエン等の揮発性芳香族炭化水素のほか、多環芳香族炭化水素(PAHs)が含まれます。その多くはアルキル側鎖を持ちます。これらの炭化水素は微生物によって生分解を受けます。好気的な条件下での分解は、嫌気的条件下での分解よりも圧倒的に速い速度で起こります。この分解の初発反応は、酸素添加酵素(オキシゲナーゼ)による水酸基の付与です。アルカン類の場合は、基質に水酸基を一つ導入するモノオキシゲナーゼによって、メチル基がアルコールに変わります。一方、芳香族炭化水素の場合は、基質に二つの水酸基を導入するジオキシゲナーゼによって、芳香環に二つの水酸基が導入されます。
【図6】背景:原油成分の微生物分解
本調査では、以下の三つの分離法を用い、インドネシア海水から石油分解菌の分離を試みました。
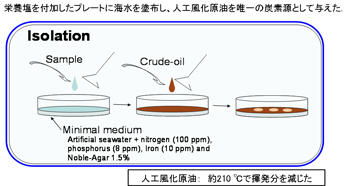
a. 自然海水からの石油分解菌の直接分離
人工海水をベースにした寒天培地に、唯一の炭素源として人工風化アラビアンライト原油を塗布したものを用いて、自然海水からの集積培養を行わずに、石油分解菌の分離培養を30℃で行いました。生じたコロニーは、同じ寒天培地上で画線培養し、均一なコロニーを形成するまで画線培養を繰り返し、純化しました(【図7】)。
【図7】自然海水からの石油分解菌の直接分離
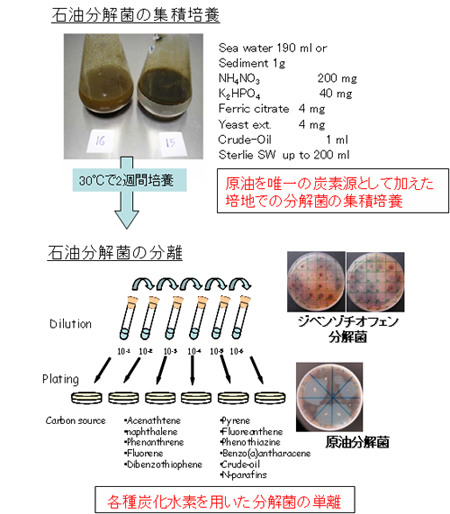
b. 石油分解菌バッチ培養からの分離
インドネシア海水に栄養塩溶液と原油を加え、30℃で2週間程度培養した後、その培養液から石油分解菌を分離しました。炭素源としては、前項a.と同様に原油を用いました。PAH分解細菌の分離には、多環芳香族化合物(PAHs/PACs:ナフタレン、ジベンゾチオフェン、フェナントレン、ピレン等)をsublimation法(Appl Environ Microbiol. 66: 439-442. 2000)によって寒天培地表面に加えたものも用いました。この寒天培地を30℃で培養し、生育、クリアゾーンの形成及び培地の着色によって分解細菌の判定を行いました(【図8】)。
【図8】石油分解菌バッチ培養からの分離
c. 石油分解菌連続培養からの分離
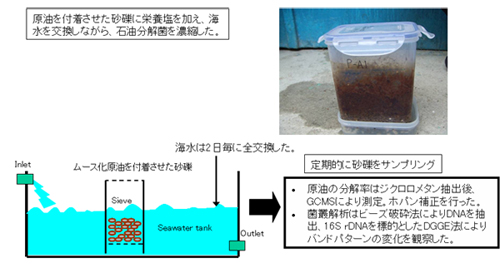
閉鎖系であるバッチ培養では、開放系である自然環境中の原油汚染事故で集積されるような石油分解菌を分離できないおそれがあるので、原油を付着させた砂れきに石油分解菌が集積するような連続培養法も併用して、石油分解菌を分離しました(【図9】)。
【図9】石油分解菌連続培養からの分離
上記三つの分離方法を用いて、インドネシア海水から1,500株以上の石油分解菌及びPAH分解細菌を分離しました。分離した株の16S rRNA遺伝子配列を決定し、BLAST検索等を用いて系統解析を行いました。
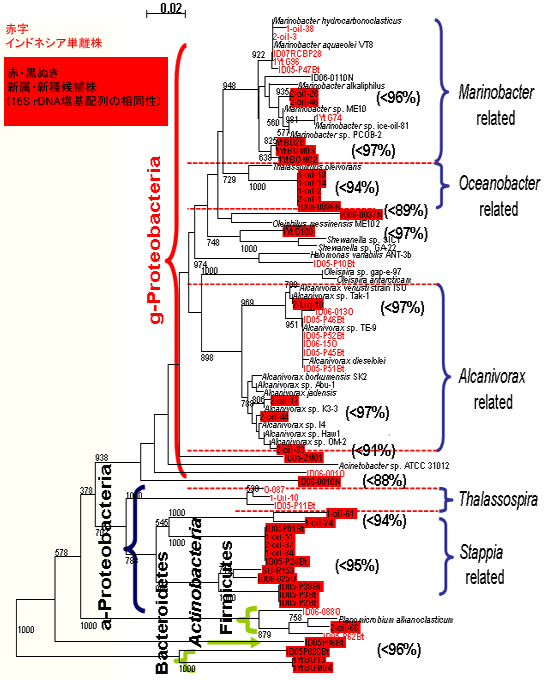
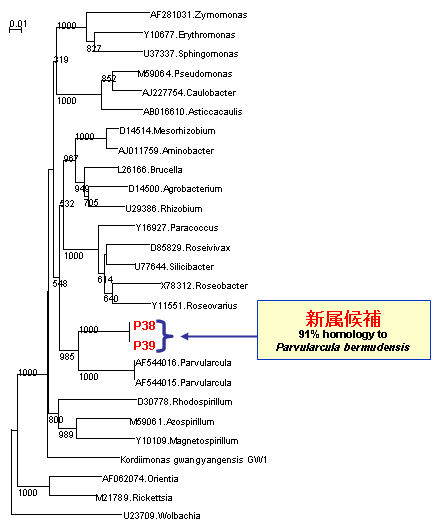
石油分解菌として、既知のアルカン分解細菌Alcanivorax属、Marinobacter属が分離されました。それ以外に、これまでアルカン分解細菌として報告のないOceanobacter属に類縁の菌株が分離されました。さらに、Alphaproteobacteria 、Actinobacteria 、Flavobacterium等、多様な石油分解菌が存在することが明らかになりました。それらの多くは、新属又は新種の候補株でした(【図10】)。
【図10】インドネシアから分離したアルカン分解菌の多様性
PAH分解細菌の分離には、主にフェナントレンとジベンゾチオフェンを炭素源として用いましたが、ジベンゾチオフェン分解細菌に比べ、フェナントレン分解細菌が数多く単離されました。系統解析の結果、PAH分解細菌の多くはAlphaproteobacteriaに属していました(【図11】)。
【図11】インドネシアから分離した芳香族炭化水素分解菌の多様性
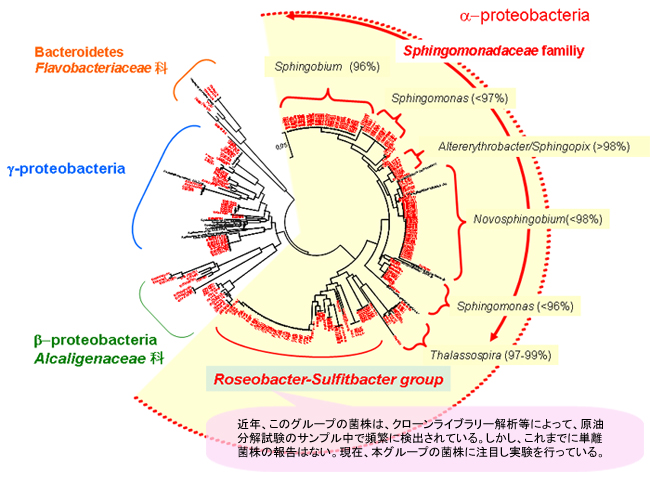
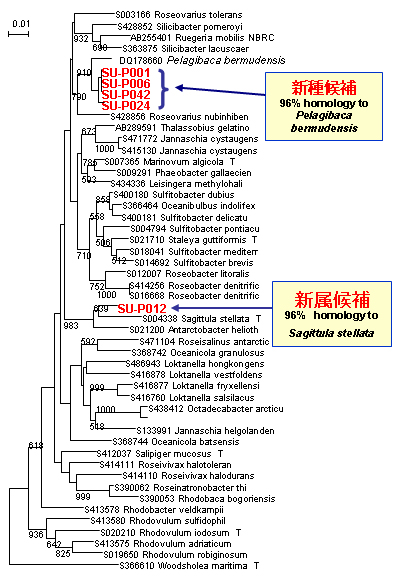
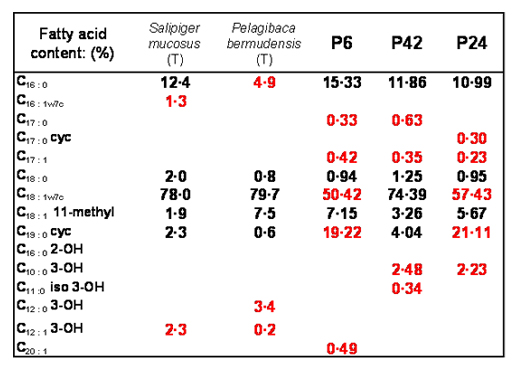
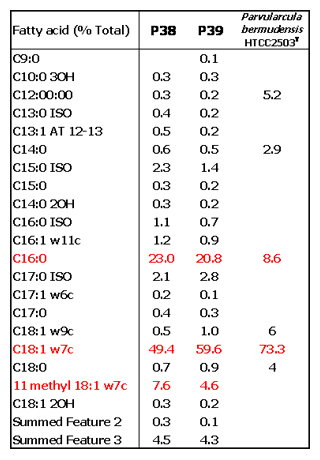
これまでに、海洋のPAH分解細菌ではCycloclasticus属について数多くの報告があります。しかし、本調査の結果では、インドネシア海水中から多数の新属・新種候補株を含むPAH分解細菌が確認されました。特に、Alphaproteobacteria綱のRoseobacter-Sulfitobacter属類縁細菌は、これまでに分子生物学的手法により海水中での原油の分解に重要な働きを持つことが示唆されていますが、分解細菌として分離されていませんでした。また、Alphaproteobacteria以外にもGammaproteobacteriaやこれまでに海洋での報告がないBetaproteobacteria、Bacteroidetes等、多様な細菌種の存在が明らかになりました(【図12】)。
以上、インドネシア海水からの石油分解菌から多様な石油分解菌を分離し、以下のようにまとめることができます。
- インドネシア海水から、約1,500株の細菌を分離しました。
- 明らかに重複のない691株を、日本に移転しました。
- 分離株の系統解析の結果、新種(属)候補186株、石油分解菌として報告のない53属を含む、多様な石油分解菌を分離・保存しました。
- 実施概要
- 石油分解菌の検出とその多様性
- 石油分解遺伝子の多様性
- 石油分解菌の特性の解明
- 石油分解菌を活性化する条件の検討
- 収集した菌株の提供
お問い合わせ
- 独立行政法人製品評価技術基盤機構 バイオテクノロジーセンター バイオ技術評価・開発課(かずさ)
-
TEL:0438-20-5764
住所:〒292-0818 千葉県木更津市かずさ鎌足2-5-8 地図
お問い合わせフォームへ









